Комплементарное взаимодействие. Комплементарное взаимодействие генов Какие гены людей обладают комплементарным действием
К комплементарным,
или дополнительно действующим, генам относятся такие неаллельные гены, которые при совместном проявлении обуславливают развитие нового признака. На примере наследования окраски цветков у душистого горошка можно понять сущность комплементарного действия генов. При скрещивании двух рас этого растения с белыми цветками у гибридов F1 цветки оказались пурпурными. При самоопылении растений из F1 в F2 наблюдалось расщепление растений по окраске цветков в отношении близком к 9:7. пурпурные цветки были обнаружены у 9/16 растений, белые у 7/16. Объяснение такого результата состоит в том, что каждый из доминантных генов не может вызвать появление окраски, определяемой пигментом антоцианом. У душистого горошка есть ген А, обусловливающий синтез бесцветного предшественника пигмента - пропигмента. Ген В определяет синтез фермента, под действием которого из пропигмента образуется пигмент. Цветки душистого горошка с генотипом ааВВ и ААbb имеют белый цвет: в первом случае есть фермент, но нет пропигмента, во втором -есть пропигмент, но нет фермента, переводящего пропиг-мент в пигмент. Проведем скрещивание двух растений ду шистого горошка с белыми цветками: У дигетерозиготных растений есть и пропигмент (А), и фермент (В), участвующие в образовании пурпурного пигмента. Формирование такого, казалось бы, элементарного признака, как окраска цветков, зависит от взаимодействия по крайней мере двух неаллельных генов, продукты которых взаимно дополняют друг друга. Такая форма взаимодействия генов разных аллельных пар носит название комплементарности - взаимодополнения.
Эпистаз.

Эпистатическое взаимодействие неаллельных генов в определенном смысле противоположно комплементарному действию генов. Сущность эпистаза сводится к подавлению проявления генов одной аллельной пары генами другой. Гены, подавляющие действие других неаллельных генов, называются супрессорами или подавителями. Они могут быть как доминантными, так и рецессивными, например А - В- или bbA - . Наследование окраски у свиней демонстрирует доминантный Эпистаз. При скрещивании черных и белых свиней из разных пород в F1 появляются белые потомки. Их скрещивание между собой приводит к появлению белых (12/16), черных (3/16) и красных (1/16) поросят. Все белые поросята имеют минимум один доминантный генподавитель I. Черные поросята гомозиготны по рецессивному аллелю i, не препятствующему формированию окраски, и несут доминантный аллель Е, детерминирующий образование черного пигмента. Красные поросята (eeii) лишены доминантного геноподавителя I и доминантного гена, определяющего черную окраску.
Полимерия.
В некоторых случаях установлено, что проявление конкретного признака зависит от количества доминантных генов, вносящих вклад в его развитие. Например, при скрещивании краснозерных пшениц с белозерными было установлено, что растения с генотипом А1А1А2А2 имеют красные зерна, растения а1а1а2а2 – белые зерна, растения с тремя доминантными генами – красноватую окраску, а растения с двумя и одним геном – более бледную окраску. Таким образом, накопление определенных аллелей в генотипе может вести к изменению выраженности признаков.
2.25Плейотропное действие генов
Это зависимость нескольких признаков от одного гена, то есть множественное действие одного гена. В дрозофилы ген белого цвета глаз одновременно влияет на цвет тела, длины, крыльев, строение полового аппарата, снижает плодовитость, уменьшает продолжительность жизни. У человека известна наследственная болезнь - арахнодактилия ("паучьи пальцы"-очень тонкие и длинные пальцы), или болезнь Марфана. Ген, отвечающий за эту болезнь, вызывает нарушение развития соединительной ткани и одновременно влияет на развитие нескольких признаков: нарушение строения хрусталика глаза, аномалии в сердечно-сосудистой системе.
Плейотропное действие гена может быть первичным и вторичным. При первичнойплейотропииген проявляет свой множественный эффект. Например, при болезни Хартнупа мутация гена приводит к нарушению всасывания аминокислоты триптофана в кишечнике и его реабсорбции в почечных канальцах. При этом поражаются одновременно мембраны эпителиальных клеток кишечника и почечных канальцев с расстройствами пищеварительной и выделительной систем.
При вторичной плейотропии есть один первичный фенотипний проявление гена, вслед за которым развивается ступенчатый процесс вторичных изменений, приводящих к множественным эффектам. Так, при серповидно клеточной анемии у гомозигот наблюдается несколько патологических признаков: анемия, увеличенная селезенка, поражение кожи, сердца, почек и мозга. Поэтому гомозиготы с геном серповидно клеточной анемии гибнут, как правило, в детском возрасте. Все эти фенотипные проявления гена составляют иерархию вторичных проявлений. Первопричиной, непосредственным фенотипним проявлением дефектного гена является аномальный гемоглобин и эритроциты серповидной формы. Вследствие этого происходят последовательно другие патологические процессы: слипание и разрушение эритроцитов, анемия, дефекты в почках, сердце, мозге - эти патологические признаки вторичны.
При плейотропии, ген, воздействуя на какой то один основнй признак, может также менять, модифицировать проявление других генов, в связи с чем введено понятие о генах-модификаторах. Последние усиливают или ослабляют развитие признаков, кодируемых "основным" геном.
Показателями зависимости функционирования наследственных задатков от характеристик генотипа является пенетрантность и экспрессивность.
Рассматривая действие генов, их аллелей необходимо учитывать и модифицирующее влияние среды, в которой розвивается организм. Если растения примулы скрещивать при температуре 15-20 ° С, то в F1 согласно менделивской схеме, все поколения будут иметь розовые цветы. Но когда такое скрещивание проводить при температуре 35 °С, то все гибриды будут иметь цветы белого цвета. Если же осуществлять скрещивания при температуре около 30 ° С, то возникает разное соотношение (от 3:1 до 100%) растений с белыми цветами.
Такое колебание классов при расщеплении в зависимости от условий среды получило название пенетрантность - сила фенотипного проявления. Итак, пенетрантность- это частота проявления гена, явление появления или отсутствия признака у организмов, одинаковых по генотипу.
Пенетрантность значительно колеблется как среди доминантных, так и среди рецессивных генов. Наряду с генами, фенотип которых появляется только при сочетании определенных условий и достаточно редких внешних условий (высокая пенетрантность), у человека есть гены, фенотипное проявление которых происходит при любых соединениях внешних условий (низкая пенетрантность). Пенетрантностью измеряется процентом организмов с фенотипным признаком от общего количества обследованных носителей соответствующих аллелей.
Если ген полностью, независимо от окружающей среды, определяет фенотипное проявление, то он имеет пенетрантность 100 процентов. Однако некоторые доминантные гены проявляются менее регулярно. Так, полидактилия имеет четкое вертикальное наследования, но бывают пропуски поколений. Доминантная аномалия - преждевременное половое созревание - присуще только мужчинам, однако иногда может передаться заболевания от человека, который не страдал этой патологией. Пенетрантностью указывает, в каком проценте носителей гена оказывается соответствующий фенотип. Итак, пенетрантность зависит от генов, от среды, от того и другого. Таким образом, это не константное свойство гена, а функция генов в конкретных условиях среды.
Экспрессивность(лат. ехргеssio - выражение) - это изменение количественного проявления признака в разных особей-носителей соответствующего аллелей.
При доминантных наследственных заболеваниях экспрессивность может колебаться. В одной и той же рсемье могут проявляться наследственные болезни от легких, едва заметных до тяжелых: различные формы гипертонии, шизофрении, сахарного диабета и т.д. Рецессивные наследственные заболевания в пределах семьи проявляются однотипно и имеют незначительные колебанийния экспрессивности.
В том случае, когда признак появляется только при сочетании двух доминантных аллелей разных генов (например, А и В) их взаимодействие называют комплементарностью , а сами гены комплементарными (дополняющими друг друга). При этом каждый из взаимодействующих неаллельных генов в отсутствии другого не обеспечивает формирования признака. Комплементарное взаимодействие двух генов можно обозначить в виде формулы: Аа Вв
Известным примером комплементарного взаимодействия неаллельных генов является наследование окраски цветков душистого горошка (Lathyrus odoratus) при скрещивании двух родительских форм с белыми цветками ААвв и ааВВ. В потомстве F1 (АаВв), а также в F2 (фенотипический класс А-В-) будет появляться новая окраска – пурпурная.
При этом в F2 соотношение классов с окрашенными цветками (А-В-) и классов с неокрашенными цветками (А- bb; ааВ- и ааbb) будет соответствовать формуле 9:7 . Основными пигментами, определяющими окраску цветков душистого горошка, являются антоцианы.
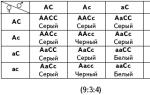
Аналогичным примером является образование коричневого пигмента у шелкопрядов. Известно, что синтез пигмента ксантомматина (пигмент оммохромового ряда) осуществляется из триптофана.У шелкопряда известны рецессивные мутации двух неаллельных генов, которые, будучи в гомозиготном состоянии (генотипы ааВВ или ААbb) делают насекомых неокрашенными, поскольку мутации в любом из генов А или В блокирует синтез пигмента, а промежуточные соединения L-кинуренин и 3-оксикинуренин не имеют окраски. У гибридов первого поколения (АаВb) синтез пигмента восстанавливается в результате комплементарного взаимодействия генов А и В. В F2 наблюдается расщепление 9:7. По такому же принципу наследуется содержание цианида у растений клевера. У земляники развитие «усов», т. е. вегетативных самоукореняющихся побегов, определяется доминантным аллелем, а «безусость» – рецессивным. Но существуют такие формы безусой земляники, которые при скрещивании друг с другом дают гибриды F1 с сильно выраженным признаком «усатости». Было показано, что в потомстве такого гибрида в F2 наблюдается расщепление, близкое к отношению 9:7. Это наиболее простые примеры комплементарного взаимодействия неаллельных генов, когда действие каждого из них в отдельности вообще не проявляется. Признак развивается лишь в результате взаимодействия доминантных аллелей двух неаллельных генов. В силу этого в F2 обнаруживается только два фенотипических класса в соотношении 9:7. Известны, однако, случаи, когда один или оба комплементарных гена характеризуются самостоятельным проявлением . В соответствии с этим меняется и характер расщепления в F2. Пример комплементарного действия генов при наследовании формы плода у тыквы (Cucurbita pepo). В связи с тем, что генотипы ААвв и ааВВ фенотипически не различимы, то они в сумме дают цифру 6. Дисковидная форма возникает в результате взаимодействия двух доминантных генов (А и В), а удлиненная форма плода – как следствие сочетания их рецессивных аллелей. Рис. 33. Наследование формы плода у тыквы при комплементарном взаимодействии двух генов (расщепление 9: 6: 1)

ааВВ ААвв сферической формы формы F1: АаВв дисковидной формы F2: 9 А-В- : 3 ааВ- : 3 А-вв: аавв дисковидная сферическая удлиненная. Расщепление 9: 3: 4.
В том случае, когда фенотип одного из родителей (например, имеющего генотип ааВ-) совпадает с фенотипом рецессивной гомозиготы (ааbb), расщепление в F2 будет 9: 3: 4.
Наследование формы гребня у кур. В данном примере каждый из комплементарных доминантных генов характеризуется собственным специфическим эффектом, а взаимодействие между ними приводит к новообразованию, к новому выражению признака. Расщепление в F2 по фенотипу при этом полностью соответствует менделевскому отношению 9:3:3:1, ибо каждый из четырех классов (А-В-, А-bb, ааВ-, aabb) имеет свой особый фенотип. Откуда появляется дикий тип при скрещивании мутантных форм? Значит признаки комплементируют между собой (дополняют друг друга). К комплементарным, или дополнительным генам относят такие гены, которые при совместном действии в генотипе в гомо- или гетерозиготном состояниях (А-В-) обусловливают развитие нового признака.
Действие же каждого гена в отдельности (А-bb или ааВ-) воспроизводит признак лишь одного из скрещиваемых родителей.
Проявление признаков организма зависит не только от унаследованных генов, но и от того, как гены взаимодействуют между собой. Генотип содержит информацию об определённых признаках, но они могут не проявляться в фенотипе, а также проявляться различным образом, в зависимости от того, как взаимодействуют гены.
Аллельное взаимодействие
Каждая хромосома имеет гомологичную хромосому, полученную от другого родителя. В этих хромосомах симметрично расположены аллельные гены, определяющие альтернативные признаки.

Рис. 1. Гомологичные хромосомы.
Как в фенотипе проявится тот или иной унаследованный признак - зависит от типа взаимодействия генов.
Доминирование
Доминирование бывает полным и неполным.
В случае полного доминирования проявляется признак аллельного гена, называемого доминантным (А).
Альтернативный признак называется рецессивным (а) и проявляется только при отсутствии доминантного.
ТОП-3 статьи которые читают вместе с этой
При неполном доминировании появляется новый, промежуточный признак. Например, у некоторых растений красный цвет (А) лепестков доминирует над белым (а).
Если при полном доминировании лепестки либо красные (АА и Аа), либо белые (аа), то при неполном гетерозигота Аа будет иметь розовые лепестки.
Кодоминирование
При наследовании 4 группы крови работает принцип кодоминирования - когда аллельные гены Iᵇ и Iᵃ действуют совместно и ни один не является доминантным или рецессивным.
Сверхдоминирование
Если у гетерозигот признак проявляется сильней, чем у любой гомозиготы, то такой тип генного взаимодействия называют сверхдоминированием.
АА ˂ Аа ˃ аа
К примеру, у дрозофилы есть гены, определяющие продолжительность жизни. Признак проявляется таким образом:
- аа - нежизнеспособные особи;
- АА - особи с нормальной продолжительностью жизни;
- Аа - повышенная продолжительность жизни.
Множественный аллелизм
В некоторых популяциях признаки кодируются не парой аллельных генов, а несколькими аллелями, возникшими в результате мутаций. Таких аллелей может быть несколько десятков.
При этом возможны разные типы взаимодействия генов. Гены могут находиться в отношениях полного и неполного доминирования.
С ˃ сᵃ ˃ сᵇ ˃ с
Ген С доминирует над любым геном, ген сᵃ доминирует над всеми, кроме С и т. д. Ген с проявляется только в гомозиготном состоянии (сс).

Рис. 2. Множественный аллелизм у кроликов.
Неаллельное взаимодействие
Неаллельные гены также влияют друг на друга.
Примерами таких воздействий является:
- плейотропия;
- эпистаз;
- полимерия;
- комплементарность.
Плейотропное действие заключается во влиянии одного гена на несколько признаков. Например, у душистого горошка один и тот же ген определяет:
- пурпурную окраску лепестков;
- пигментацию прилистников;
- тёмную окраску плодов.
Плейотропный эффект широко распространён в природе.
Эпистатическое взаимодействие - это подавление генов одной аллельной пары генами другой аллельной пары.
Оно бывает:
- доминантное (А ˃ В);
- рецессивное (аа ˃ В).
При полимерном наследовании несколько неаллельных генов контролируют один признак, причём степень его выраженности может зависеть от числа доминантных генов (кумулятивный эффект), либо не зависеть.
Комплементарное взаимодействие называют также дополнительным, т. к. при нём неаллельные гены совместно определяют признак. Это может происходить даже если один из них или оба по отдельности не кодируют такой признак.
К комплементарным , или дополнительным , генам относят такие гены, которые при совместном действии в генотипе в гомо- или гетерозиготном состояниях (А-В-) обусловливают развитие нового признака.
Действие же каждого гена в отдельности (А-bb или ааВ-) воспроизводит признак лишь одного из скрещиваемых родителей.
Впервые такого рода взаимодействие было обнаружено у душистого горошка (Lathyrus odoratus). При скрещивании двух рас этого растения с белыми цветками у гибрида F 1 цветки оказались пурпурными. При самоопылении растений F 1 в F 2 наблюдалось расщепление по окраске цветков в отношении, близком к 9:7. Один фенотипический класс (9 / 16) имел такую же окраску цветков, как и у растений первого поколения, а второй (7 / 16) - белую окраску, такую же, как у родительских растений.
Чтобы выяснить, укладывается ли это расщепление в схему дигибридного менделевского расщепления, представим себе, что у каждой исходной расы душистого горошка имеется в гомозиготном состоянии лишь по одной из доминантных аллелей (AAbb и ааВВ), которые при взаимодействии определяют развитие окраски. Поскольку у гибрида первого поколения присутствуют доминантные аллели обоих генов (АаВb), цветки гибридных растений F 1 будут окрашенными. Во втором поколении происходит расщепление в отношении 9 / 16 А-В-: 3 / 16 А-bb: 3 / 16 ааВ-: 1 / 16 aabb. Каждый из генов в отдельности не может обусловить развитие окраски, так как выработка антоциановых пигментов осуществляется лишь при наличии доминантных аллелей обоих генов. Поэтому растения с генотипами А-bb, ааВ- и aabb имеют белые цветки и во втором поколении наблюдается расщепление по фенотипу в отношении 9: 7. Анализирующим скрещиванием и анализом в F 3 можно точно подтвердить данное выше объяснение.
Приведем еще несколько примеров, иллюстрирующих действие комплементарных генов у растений и животных.
У земляники развитие «усов», т. е. вегетативных самоукореняющихся побегов, определяется доминантной аллелью, а «безусость» - рецессивной. Но существуют такие формы безусой земляники, которые при скрещивании друг с другом дают гибрид F 1 с сильно выраженным признаком «усатости». Исследованиями Т. С. Фадеевой было показано, что в потомстве такого гибрида в F 2 получается расщепление, близкое к отношению 9: 7, а именно: из 752 растений F 2 419 оказались с усами, 333 - без усов. Это соответствует теоретически ожидаемому расщеплению: 752 X 9 / 16 = 423 и 752 X 7 / 16 = 329.
У белого клевера имеются формы с высоким и низким содержанием цианида. Цианиды, как известно, блокируют дыхательный фермент, но повышают активность папаина (растительной протеазы), катепсина и других ферментов. Высокое содержание цианида в белом клевере связано с усиленным вегетативным ростом без снижения его кормовых качеств. При скрещивании растений с высоким и низким содержанием цианида в F 1 доминирует первое свойство, а в F 2 наблюдается расщепление, близкое к отношению 3: 1.
Эти результаты указывают на то, что в данном случае альтернативные признаки определяются одной парой аллелей. Но иногда при скрещивании двух растений клевера с низким содержанием цианида гибриды F 1 характеризуются высоким его содержанием, а в F 2 расщепление оказывается близким к отношению: 9 / 16 - с высоким содержанием цианида и 7 / 16 - с низким. Так же, как у душистого горошка, в данном случае имеет место обычное дигибридное расщепление, в котором 9 / 16 потомков обладают двумя доминантными генами А-В-, а 7 / 16 относятся к трем остальным фенотипически неотличимым классам: 3 / l 6 A-bb + 3 / 16 ааВ + 1 / 16 aabb = 7 / 16 . Доминантные аллели разных генов в отдельности не увеличивают содержание цианида по сравнению с тем низким уровнем, который характерен для растения, гомозиготного по рецессивным аллелям обоих генов, но при совместном действии доминантных аллелей обоих генов содержание цианида повышается.
Подобное явление можно показать на примере кукурузы. При скрещивании некоторых форм кукурузы с белыми зернами в F 1 зерна в початках оказываются пурпурными. В F 2 происходит расщепление на 9 / 16 пурпурных (А-В-) и 7 / 16 белых (ааВ-, А-bb и aabb).
До сих пор мы рассматривали примеры комплементарного взаимодействия доминантных генов, при котором каждый из генов в отдельности не обладал способностью вызывать развитие признака. Последний развивался лишь в результате взаимодействия доминантных аллелей двух генов. В силу этого в F 2 обнаруживались только два фенотипических класса в соотношении 9:7. Известны, однако, случаи, когда один или оба комплементарных гена характеризуются самостоятельным проявлением. В соответствии с этим меняется и характер расщепления в F 2 .
Рассмотрим наследование трех типов окраски шерсти у мышей: дикой, или рыжевато-серой (агути), черной и белой. Окраска дикого типа зависит от наличия гена, определяющего развитие окраски, и от гена, обусловливающего распределение пигмента по длине волоса. Каждый волос у мышей агути имеет по длине кольцо желтого пигмента, а в основании и на конце волоска - черный пигмент. Такое зонарное распределение пигментов и создает окраску агути, свойственную диким грызунам (белка, кролик, морская свинка и др.). У черных мышей отсутствует зонарное распределение пигмента - волосы до всей длине окрашены равномерно. Белые мыши с красной радужной оболочкой глаз, так называемые альбиносы, лишены пигмента.
Надо сказать, что альбинизм встречается у животных почти всех классов - млекопитающих, птиц, амфибий и др. Встречается альбинизм и у человека. Так, например, иногда у родителей-негров рождаются дети альбиносы, т. е. с белой кожей и белыми волосами, но с чертами лица негритянского типа. Известны случаи, когда в семье негров рождаются двойни (разнояйцевые), и один из детей оказывается альбиносом. Такой ребенок имеет рецессивную аллель гена альбинизма в гомозиготном состоянии.
Окраска шерсти у мышей типа агути доминирует над черной, и над белой. При скрещивании черных мышей с белыми с белыми, все гибриды F 1 оказываются агути, а в F 2 наблюдается расщепление в отношении 9 / 16 агути: 3 / 16 черных: 4 / 16 белых.
Взятые в скрещивание мыши-альбиносы являются, очевидно, гомозиготными по рецессивной аллели гена окраски и доминантной аллели гена попарного распределения пигмента (ааВB), а черные мыши - гомозиготными по доминантной аллели гена окраски и рецессивной аллели гена распределения пигмента в волоске (AAbb). У гибридов F 1 (АаВb) вследствие взаимодействия доминантных аллелей обоих генов развивается окраска типа агути. Такая же окраска характерна и для 9 / 16 особей в F 2 с генотипом А-В-. Черными в F. оказываются мыши, имеющие генотип А-bb, а белыми - все остальные - (ааВ- и aabb) в силу отсутствия у них гена А, определяющего образование пигмента. Ген В в отсутствие гена А не имеет собственного проявления.
Подобные примеры наследования известны и у растений (лук, кукуруза и др.). У лука скрещивание формы, имеющей неокрашенную (белую) луковицу, с формой, имеющей желтую луковицу, дает в F 1 растения с красными луковицами, а в F 2 появляются растении с красными (9 / 16), желтыми (3 / 16) и белыми (4 / 16) луковицами. В этом случае опять-таки одна из доминантных аллелей двух генов способна действовать самостоятельно (определяет желтую окраску луковицы), а другой ген проявляется лишь в присутствии комплементарного гена.
Известны и такие случаи, когда каждый из двух комплементарных генов способен проявлять свое действие самостоятельно. Один таких примеров мы уже рассматривали при анализе наследования формы гребня у кур. Каждая из доминантных аллелей генов обусловливала развитие гребня определенной формы (гороховидной или розовидной), а взаимодействие этих генов определяло развитие новой формы гребня ореховидной. В данном примере каждый из комплементарных доминантных генов характеризуется собственным специфическим эффектом, а взаимодействие между ними приводит к новообразованию, к новому выражению признака.
Ряд подобных примеров наследования известен и у других животных и растений. Так, у дрозофилы рецессивная аллель гена scarlet в гомозиготном состоянии определяет ярко-красную окраску глаз, а рецессивная аллель другого гена - brown (также в гомозиготном состоянии) определяет коричневую окраску глаз. При скрещивании гибриды F 1 оказываются красноглазыми (дикого типа), рели же оба эти рецессивных гена находятся в гомозиготном состоянии, то такая особь оказывается белоглазой. Если скрестить красноглазых мух F 1 друг с другом, то во втором поколении по признаку окраски глаз будет наблюдаться расщепление на 4 фенотипических класса в отношении 9 / 16 красных: 3 / 16 ярко-красных: 3 / 16 коричневых: 1 / 16 белых. Такое поведение признаков в наследовании также говорит о расщеплении по двум комплементарным генам с самостоятельным действием.

Если генотип мух с коричневыми глазами условно обозначить ААbb, с ярко-красными - ааВВ, а генотип красноглазых гибридов F 1 - АаВb и белоглазых мух - aabb, то фенотипические радикалы полученных в F 2 классов могут быть представлены как А-В- (9 / 16), ааВ- (3 / 16), А-bb(3 / 16) и aabb(1 / 16).
Природа взаимодействия генов в этом случае более ясна, чем в случае наследования формы гребней у кур. Нормальная красная окраска глаз у мух обеспечивается в основном тремя видами пигментов красным, коричневым и желтым. В гомозиготном состоянии рецессивный ген а блокирует образование коричневого пигмента, вследствие чего развиваются ярко-красные глаза, а другой рецессивный ген b в гомозиготном состоянии блокирует одновременно образование красного и желтого пигментов, и поэтому развиваются коричневые глаза. В F 1 объединяются доминантные аллели этих генов, поэтому образуются все пигменты, дающие в совокупности красную окраску глаз. Новый класс белоглазых мух, появляющихся в F 2 , очевидно, является результатом одновременного блокирования синтеза всех трех пигментов.
Подобные примеры можно привести и на растительных объектах. Известно, что окраска плодов у томатов обусловливается каротиновыми пигментами (ликопины и бета-каротин), имеющими огромное значение в синтезе витаминов. Анализ наследования окраски плодов у томатов показывает, что красная окраска плодов определяется взаимодействием комплементарных доминантных генов R и Т, оранжевые плоды образуются на растениях с генотипом R-tt, желтые - на растениях с генотипом rrТ-, промежуточные, желтооранжевые - на растениях rrtt. Здесь также расщепление в F 2 соответствует генетической формуле дигибридного скрещивания 9:3:3: 1. При этом установлено, что красные и оранжевые плоды содержат наибольшее количество каротинов, а желтые - наименьшее. Двойной рецессив содержит промежуточное количество каротинов в плоде. Качественные различия в наборе каротинов соответствуют определенным различиям в генотипе.
Во всех разобранных примерах мы имели дело с комплементарным взаимодействием доминантных и рецессивных неаллельных генов. Взаимодействие доминантных генов обусловливало развитие ореховидного гребня у кур, красной окраски глаз у дрозофилы, красной окраски плодов у томатов. Взаимодействие рецессивных аллелей этих генов приводило к развитию пластинчатого, или ликвидного, гребня у кур, белых глаз у дрозофилы, желто-оранжевой окраски плодов у томатов.
Следует отметить, что в ряде случаев комплементарные гены, способные к самостоятельному проявлению, при отсутствии дополнительного гена могут давать каждый в отдельности сходный фенотипический эффект. Характер расщепления дигибрида в F 2 при этом также изменяется. Так, у тыквы (Cucurbita реро) имеются сорта с разной формой плода: сферической, дисковидной и удлиненной. Сферическая форма плода является рецессивной по отношению к дисковидной.

От скрещивания растений с плодами сферической формы, но имеющими разное происхождение, возникают гибридные растения, дающие плоды только дисковидной формы. В потомстве этих растений в F 2 появляются три фенотипических класса в отношении 9/16 дисковидными плодами, 6/16 - со сферическими и 1 / 16 - с удлиненными. Зная закономерности дигибридного расщепления при взаимодействии генов, нетрудно понять, что и здесь имеет место взаимодействие двух генов, влияющих на развитие формы плода, каждый из доминантных комплементарных генов обусловливает развитие плодов сферической формы, а их взаимодействие приводит к образованию дисковидных плодов. Взаимодействие рецессивных аллелей этих генов определяет развитие плодов удлиненной формы.
Рассматривая примеры комплементарного действия генов, мы убеждаемся, что такое взаимодействие генов приводит к развитию признаков, свойственных диким предкам данных видов (серая окраска грызунов, дисковидная форма у тыквы и т. д.). Некоторые авторы рассматривают это явление как пример атавизма. Эти представления основываются на предположении, что в процессе эволюции животных и растений доминантные гены, действующие комплементарным образом, изменились, мутировали в рецессивное состояние (А → а, В → b, С → с и т. д.).
У диких предков домашних животных и растений доминантные гены комплементарного действия поддерживались естественным отбором вместе в одном генотипе (например, серая окраска грызунов, дисковидная форма плода у тыквы, красная окраска глаз у дрозофилы и др.). При одомашнивании и проведении селекции с помощью скрещиваний и искусственного отбора комплементарные гены как бы разобщились. Генотип АаВb разлагался селекционерами на генотипы ААЬЬ и ааВВ. Поэтому при скрещивании таких организмов иногда наблюдается как бы возврат к признакам диких предков.
Мы остановились более подробно на комплементарном действии генов потому, что этот тип взаимодействия иллюстрирует один из путей возникновения комбинативной изменчивости и имеет отношение к широко используемому явлению гибридной мощности - гетерозису.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .
Комплементарность. Комплементарными (complementum – средство пополнения) называются взаимодополняющие гены, когда для формирования признака необходимо наличие нескольких неаллельных (обычно доминантных) генов. Этот тип наследования широко распространен в природе.
Комплементарное взаимодействие неаллельных генов характерно для человека, например процесс формирования половой принадлежности. Определение пола у человека происходит в момент оплодотворения, если яйцеклетку оплодотворяет сперматозоид с X - хромосомой - рождаются девочки, если с У - рождаются мальчики. Установлено, что У хромосома определяет дифференцировку половых желез по мужскому типу синтезирующих гормон тестостерон и не всегда способна обеспечить развитие организма мужского пола. Для этого нужен белок - рецептор, который синтезируется особым геном, имеющимся в другой хромосоме. Этот ген может мутировать, и тогда особь с кариотипом ХУ внешне похожа на женщину. Эти люди не могут иметь потомства, т.к. половые железы - семенники - недоразвиты, а формирование организма часто идет по женскому типу, но недоразвита матка и влагалище. Это синдром Морриса или тестикулярная феминизация.
Типичным примером комплементарности является развитие слуха у человека. Для нормального слуха в генотипе человека должны присутствовать доминантные гены из разных аллельных пар - D и Е, где D отвечает за нормальное развитие улитки, а ген Е - за развитие слухового нерва. У рецессивных гомозигот (dd) будет недоразвита улитка, а при генотипе ее - недоразвит слуховой нерв. Люди с генотипами ДДЕЕ, ДДЕе, ДдЕЕ, ДдЕе будут иметь нормальный слух, у людей с генотипами ДДее, Ддее, ддЕЕ, ддее слух будет отсутствовать.
Эпистаз -это взаимодействие неаллельных генов, противоположное комплементарному. Существует эпистатический ген или ген ингибитор, который подавляет действие как доминантного, так и рецессивного неаллельных генов. Различают доминантный и рецессивный эпистаз.
Доминантный эпистаз можно наблюдать при наследовании окраски оперения у кур.
С – синтез пигмента в пере.
с – отсутствие пигмента в пере.
J – эпистатический ген, подавляющий действие гена С.
j – не подавляет действие гена С.
Примером рецессивного эпистаза у человека может служить «бомбейский феномен» в наследовании групп крови. Он описан у женщины, получившей от матери аллель J В (третья группа крови), а фенотипически женщина имеет первую группу крови. Было выяснено, что деятельность аллеля J В подавлена редким рецессивным аллелем гена х, который в гомозиготном состоянии оказывает эпистатическое действие (I B I B хх).
Полимерия –это явление, при котором несколько доминантных неаллельных генов детерминируют (определяют) один признак. Степень проявления признака зависит от количества доминантных генов в генотипе. Чем их больше, тем интенсивнее выражен признак.
По типу полимерии наследуется цвет кожи у человека.
S 1 S 2 – темная кожа.
s 1 s 2 – светлая кожа.
Таким же образом, наследуются многие количественные и качественные признаки у человека и животных: рост, масса тела, величина артериального давления и др.
В значительной мере проявление полигенных признаков зависит также от условий окружающей среды. У человека может наблюдаться предрасположенность к различным заболеваниям: гипертонической болезни, ожирению, сахарному диабету, шизофрении и др. Данные признаки при благоприятных условиях среды могут и не проявиться или быть слабо выраженными. Это отличает полигенно наследуемые признаки от моногенных. Изменяя условия среды и проводя профилактические мероприятия можно значительно снизить частоту и степень выраженности некоторых мультифакториальных заболеваний.
Плейотропное действие гена - это детерминация одним геном нескольких признаков. Множественное действие гена обусловлено синтезом разных полипептидных цепей белка, которые влияют на развитие нескольких не связанных между собой признаков и свойств организма. Впервые это явление было обнаружено Менделем у растений с пурпурными цветками, у которых в основании черешка листьев всегда была красная окраска, а кожура семян была бурая. Эти три признака определяются действием одного гена.
Плейотропное действие гена можно наблюдать также у каракульских овец.
А – серая окраска.
а – черная окраска.
АА – серая окраска + аномалия в строении желудка (отсутствие рубца), то есть гомозиготные по доминантному гену особи погибают после рождения.
У человека плейотропное действие гена наблюдается при наследовании заболевания – синдрома Марфана . При этом один ген отвечает за наследование нескольких признаков: подвывих хрусталика глаза, аномалии в сердечно-сосудистой системе, «паучьи пальцы».
Самостоятельная работа